TD 2
Index
Function Analysis
These exercises involve plotting and describing the domain and co-domain of more complex functions. There are different ways of approaching this task, and two of such ideas will be explored here: A top-down approach for exercises "1d" and "1c" and a bottom-up approach for exercise "1g".
Exercise 1d
Initially, the behaviour of the function $f(x) = ln(2x-1)$ is unknown. As a starting point, one can think of $f$ as a composition of $g$ and $z$.
$$f(x) = g(z(x))$$
In this case, $g(z) = ln(z)$ is the "top-most" function and wraps everything else, which is neatly "packed" in $z(x) = 2x-1$. For the top-down approach, let's first focus on $g(z) = ln(z)$, whose behaviour is well known.

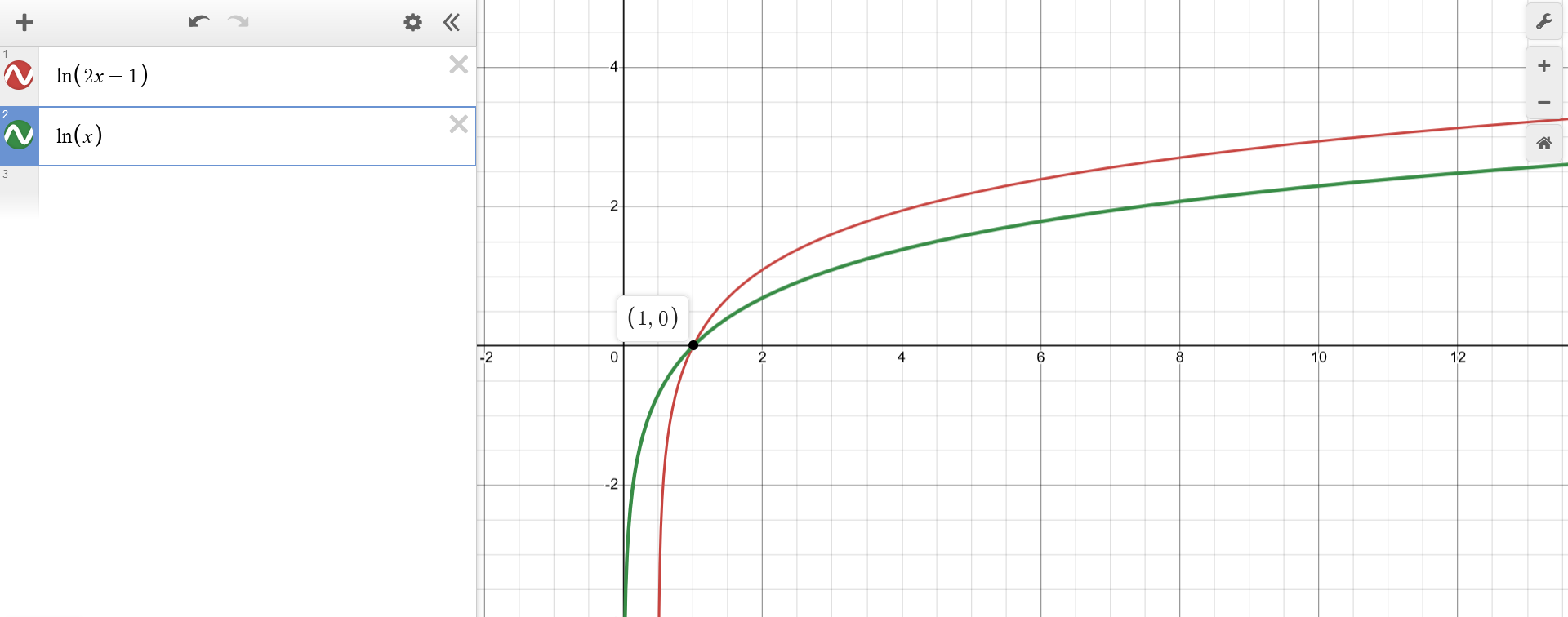
Exercise 1c
Using the top-down approach, the function $f(x) = e^{-|x-1|}$ can be first represented as $f(x) = a(b(x))$ with $a(b)=e^{-b}$. This means that $a(b)$ is just the well-known exponential function inverted along the x-axis.
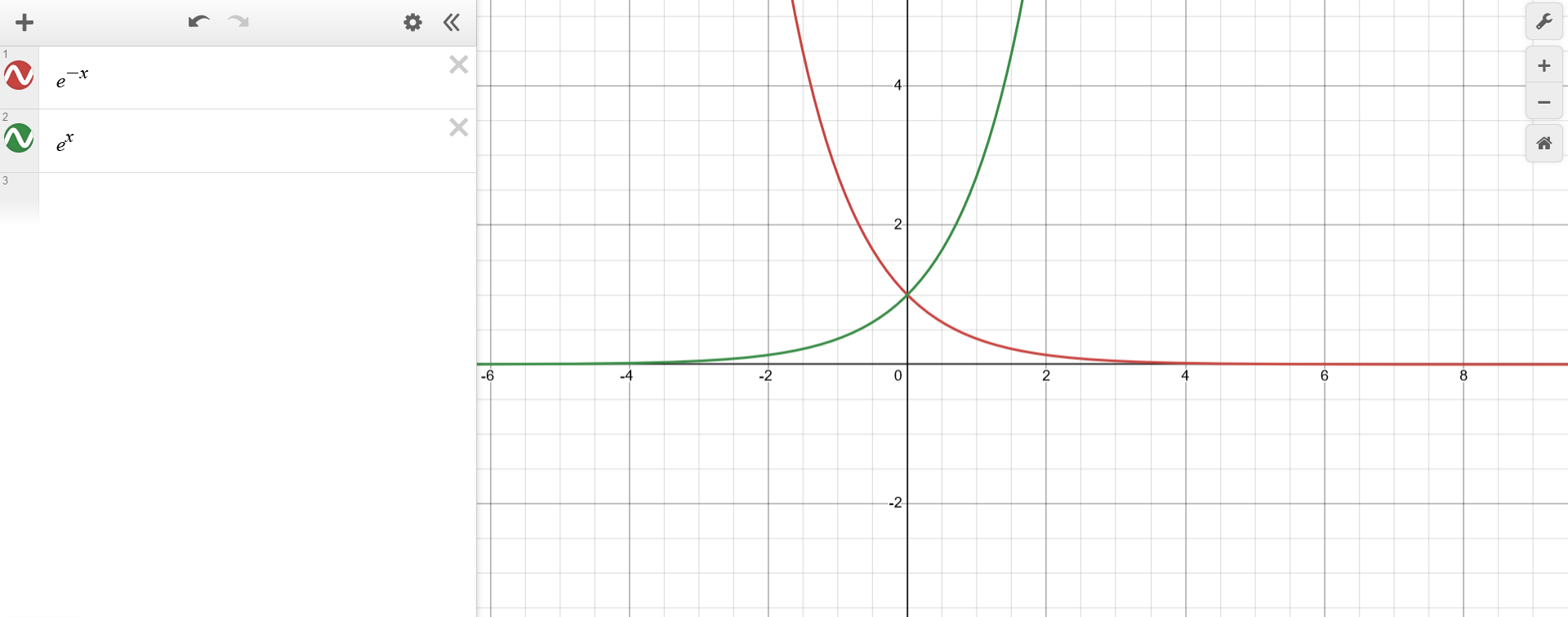
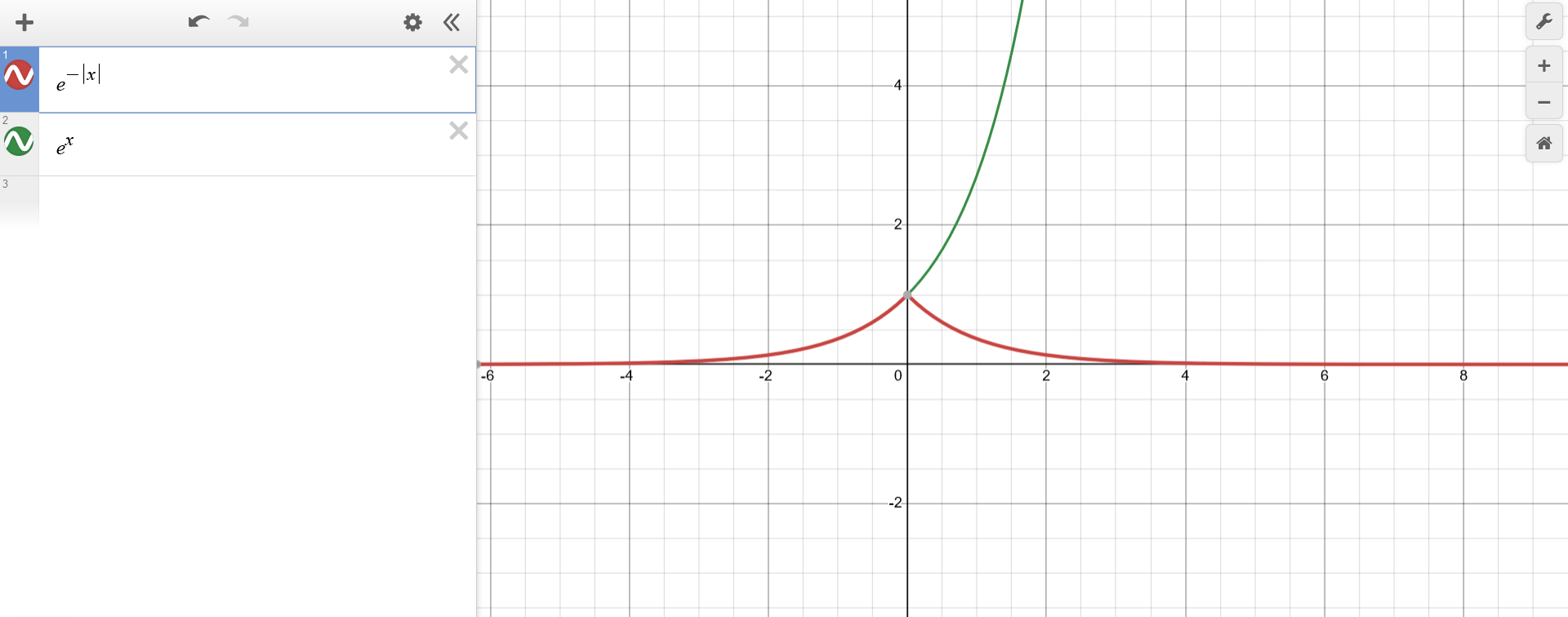
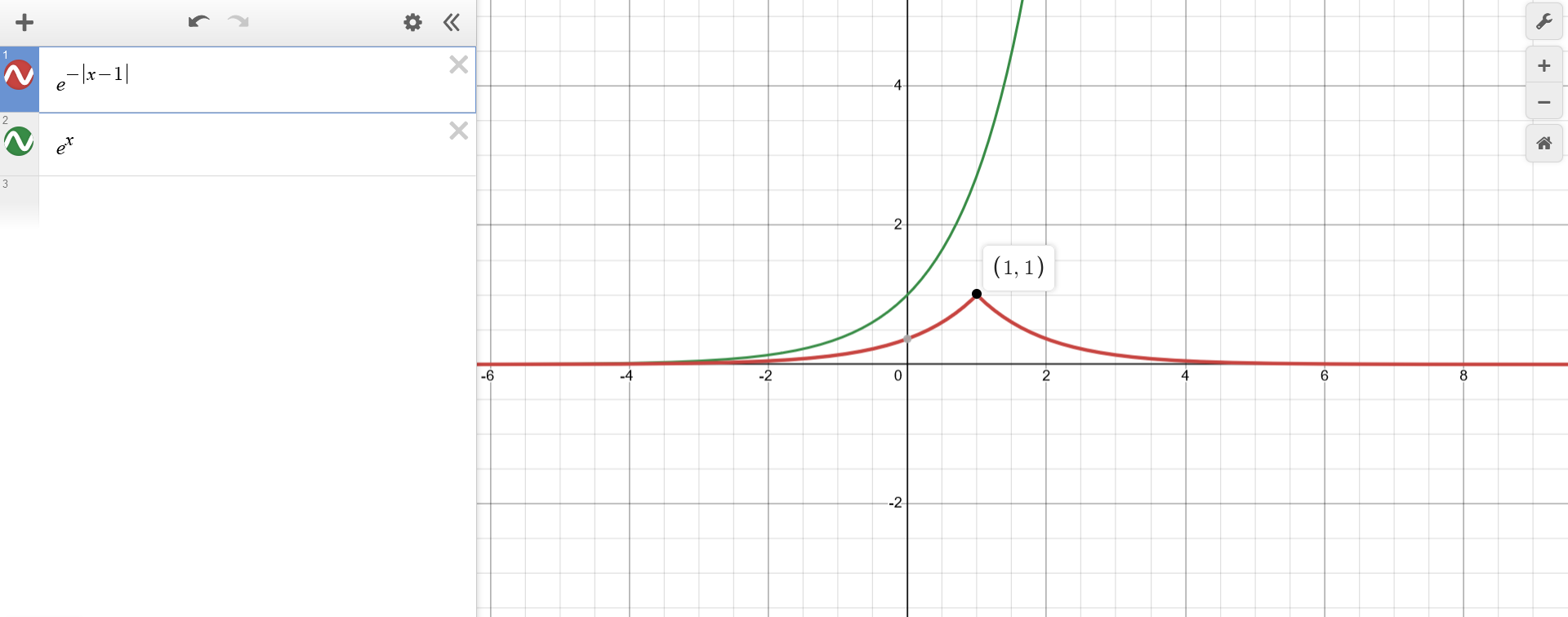
{kind=link}
Exercise 1g
The top-down approach isn't as intuitive for $f(x) = \sqrt{x^2-5x+6}$, so another strategy can be employed. Again, the function can be represented as a composition $f(x) = g(z(x))$, with $g(z) = \sqrt{z}$ and $z(x) = x^2-5x+6$. This time, $z(x)$ will be analyzed first, hence in a bottom-up fashion.
The behaviour of $z(x) = x^2-5x+6$ can be observed by plotting it, which shouldn't be problematic. It is convex (i.e. "looks upwards", as $x^2$ has a positive sign) and it can be factorized as $z(x) = (x-2)(x-3)$, which means it intersects the x-axis at $x=2$ and $x=3$.
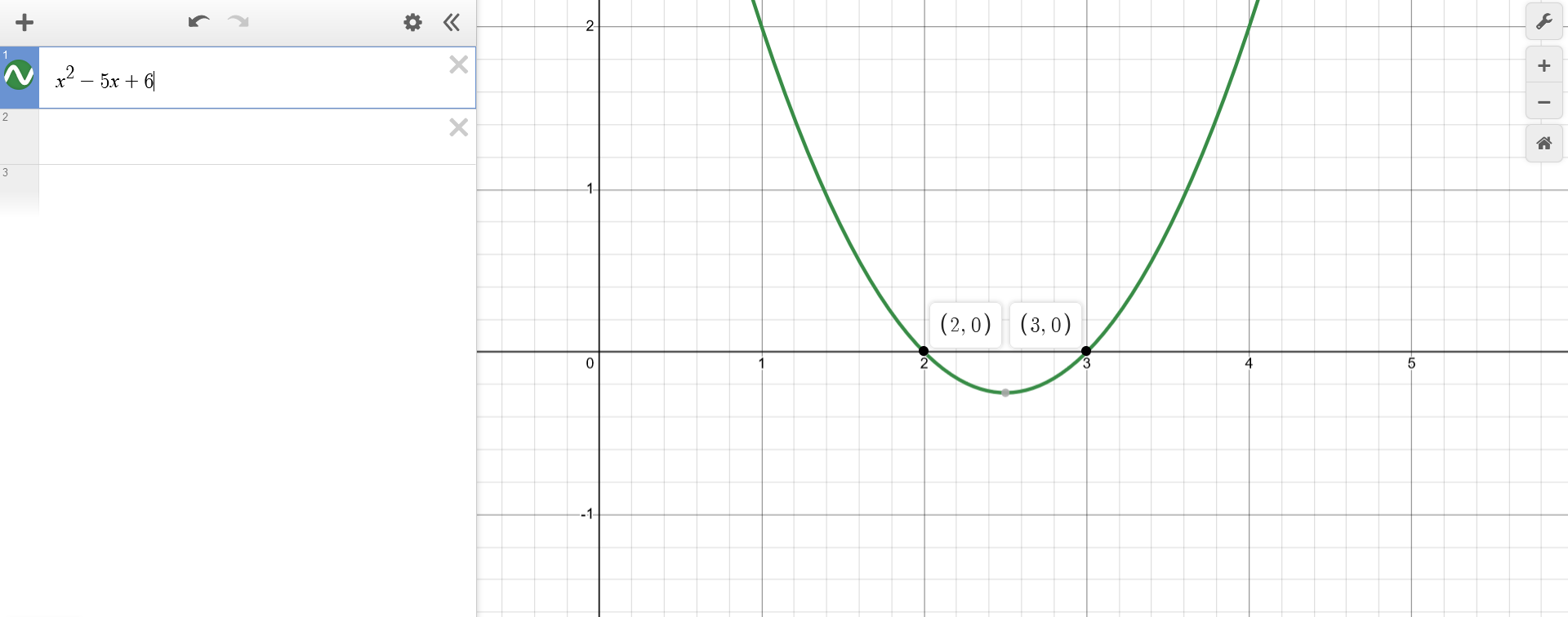

For knowing how to plot $f(x) = g(z)$ at a given point $(x^*,y^*)$, the plot of $z(x) = x^2-5x+6$ becomes handy. One can observe the value at $z^* = z(x^*)$ and use that to know how $y^* = f(z^*)$ should be, following the previous rules. After plotting $f(x)$, the domain and co-domain become once more evident.
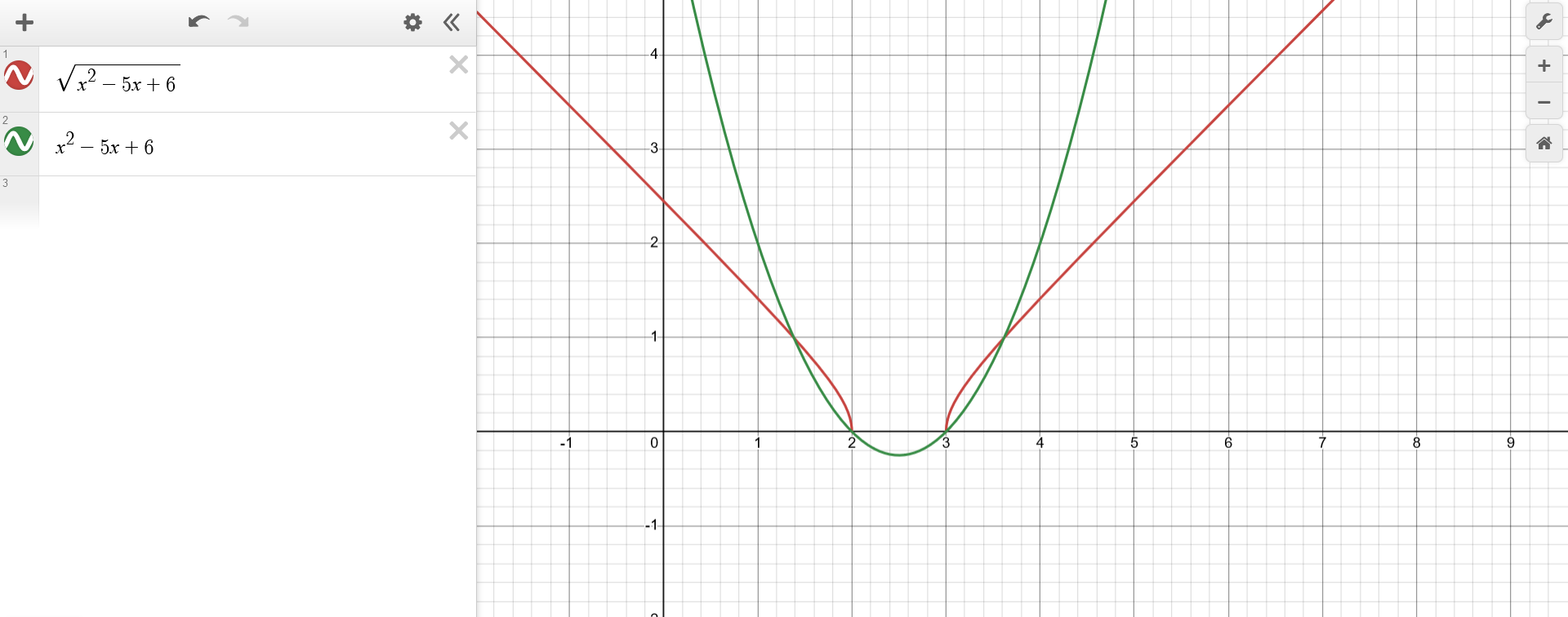
Intersection of Functions
Exercise 2a
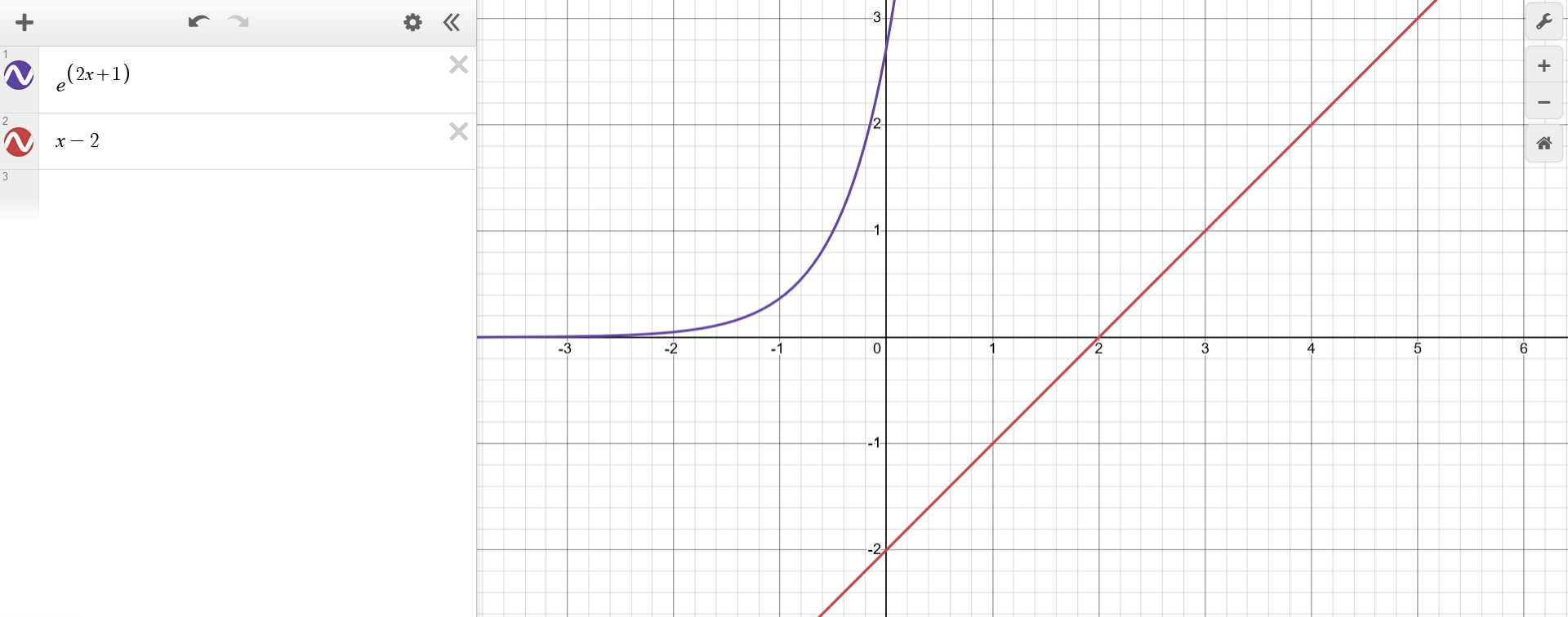
The values of $x$ that comprise the solution for the equation $e^{2x+1}=x-2$ can be inferred by plotting the functions $f(x) = e^{2x+1}$ and $g(x) = x-2$ and finding their point of intersection. After all, when two functions intersect at a given value of $x$, it means that they are equal, which is just a way of rephrasing the original problem.
In this case, one can see that $f(x) = e^{2x+1}$ is simply the exponential function shrunk by a factor of two and shifted horizontally one unit to the left. On the other hand, $g(x) = x-2$ is a simple line shifted two units down. Just by looking at $f(0)=e$ and $g(0)=-2$ and the general shape of these functions, one can plot them and see that, indeed, they do not intersect i.e. there's no solution for $e^{2x+1}=x-2$.
Problems
Exercise 3
The problem is introduced as:
In a chemical reaction, when reactants $A$ and $B$ are mixed, a species $E$ is produced. The amount of species produced depends on the amount of reactant $A$ according to the following law : $q(A) = 2ln(3A+3)$ (...) $E$ depends on $B$ linearly with a proportionality constant of $\frac{1}{2}$.Knowing this, one can define $E$ as a function of $A$ and $B$, such as $E(A,B)$. For the sake of this example, it will be assumed that the body of $E(A,B)$ simply consists of a product between two terms $q(A)$ and $p(B)$. The body of these two can be taken directly from the quote. $$\begin{aligned} E(A,B) &= q(A)p(B) \\ &= (2ln(3A+3))(\frac{B}{2}) \\ \end{aligned}$$ Now the questions of the problem can be tackled.
How much $E$ will be produced when $A = 1$?$$\begin{aligned} E(1,B) &= (2ln(3(1)+3))(\frac{B}{2}) \\ &= 2ln(3+3) \frac{B}{2} \\ &= ln(6) B \\ \end{aligned}$$
If we want to double the amount of $E$ produced, what amount of $A$ should be supplied?Given a fixed value for $A$ and $B$, let lowercase $a$ be the incognita amount required to double $E(A,B)$. $$\begin{aligned} E(a,B) &= 2E(A,B) \\ q(a)q(B) &= 2q(A)q(B) \\ ln(3a+3) &= 2ln(3A+3) \\ ln(3a+3) &= ln((3A+3)^2) \\ 3a+3 &= 9A^2+18A+9 \\ a+1 &= 3A^2+6A+3 \\ a &= 3A^2+6A+2 \\ \end{aligned}$$
Plot the curve of $E$ as a function of $B$.This is simply the plot for the identity function scaled by a factor of $\frac{1}{2}$.
If we want to double the amount of $E$, how should the amount of $B$ be changed?Given a fixed value for $A$ and $B$, let lowercase $b$ be the incognita amount required to double $E(A,B)$. $$\begin{aligned} E(A,b) &= 2E(A,B) \\ q(A)q(b) &= 2q(A)q(B) \\ \frac{b}{2} &= 2\frac{B}{2} \\ b &= 2B \\ \end{aligned}$$
Exercise 5
The problem is introduced as:
Two fish populations inhabit the same lake. The population $p_1$ consists of predators that feed on the population $p_2$. The number of fish in each population fluctuates over time according to the predator-prey relationship in a cycle : when the population $p_1$ is low, the population $p_2$ increases, and when the population $p_2$ is large, the population $p_1$ also increases because there is more food ; when the population $p_1$ is large, the population $p_2$ decreases because there are too many predators, and when the population $p_2$ is low, the population $p_1$ decreases as well because there is not enough food. Thus, both populations have a sinusoidal pattern over time.It states that both $p_1$ and $p_2$ are sinusoidal functions of time, so they could be considered as some variation of $sin(t)$. $$\begin{aligned} p_1(t) &\sim sin(t) \\ p_2(t) &\sim sin(t) \\ \end{aligned}$$
The duration of one cycle is 2 months, and there is a phase of 15 days between the phases of the two populations. (...) initially, the population $p_1$ is half that of the population $p_2$.The first statement hints at a scaling operation over the x-axis: the usual periodicity of $2\pi$ should be converted into $60$ (2 months), by plugging some factor $k$ into $sin(kx)$. Introducing $k=2\pi$ shrinks the periodicity into a value of $\frac{2\pi}{k}=\frac{2\pi}{2\pi}=1$. To extend the periodicity into $60$, $k$ must be further divided by this value. Hence, $k=\frac{2\pi}{60}=\frac{\pi}{30}$. $$\begin{aligned} p_1(t) &\sim sin(\frac{\pi}{30} t) \\ p_2(t) &\sim sin(\frac{\pi}{30} t) \\ \end{aligned}$$
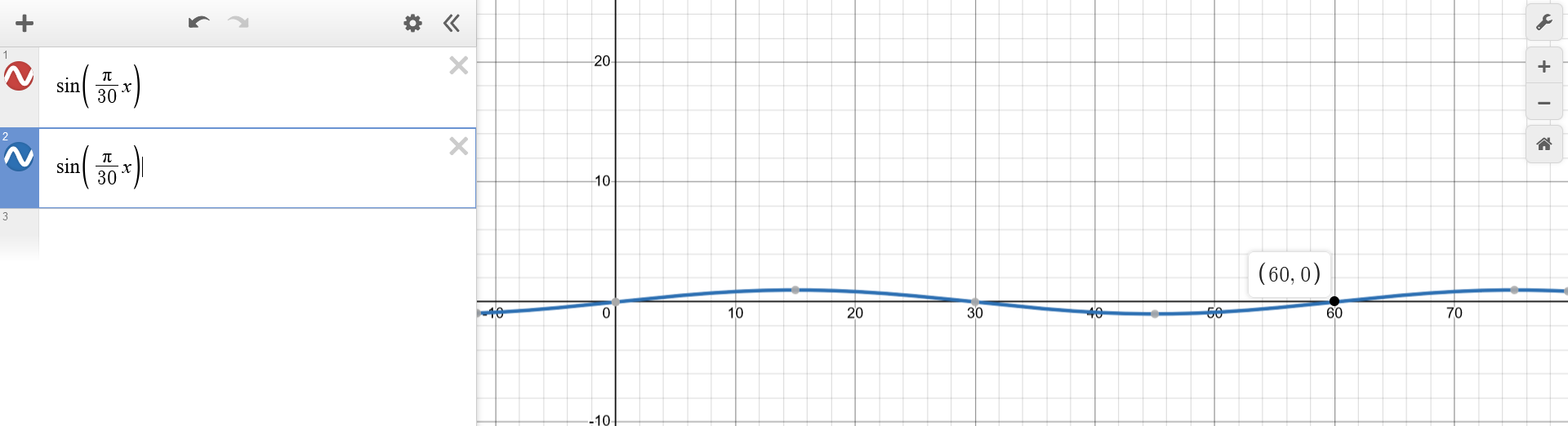
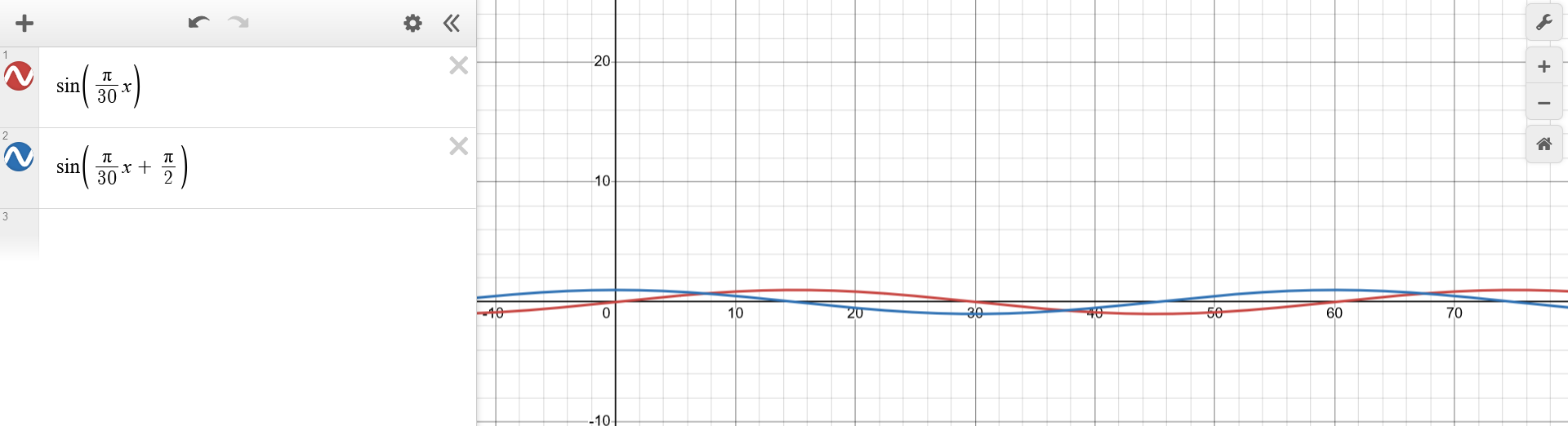
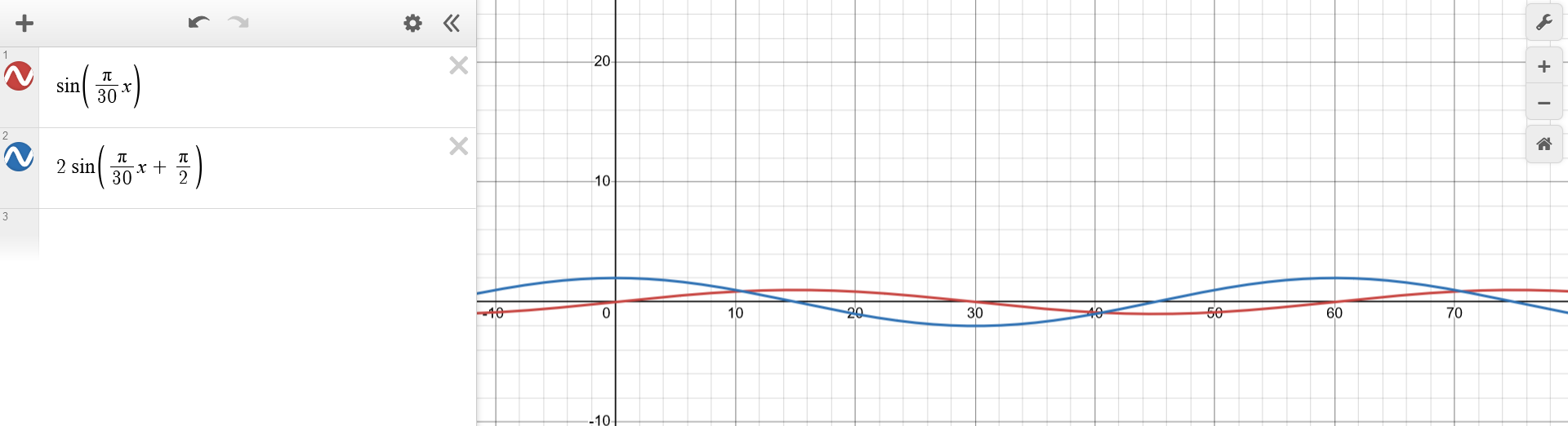
(...) knowing that the variation in the prey population is twice as large as that for the predators, (...)The final manipulation directly stated by the problem consists of performing a scaling operation over the y-axis of one of the funtions, by a factor of $2$ (or $\frac{1}{2}$). The intuitive choice is the former. $$\begin{aligned} p_1(t) &\sim sin(\frac{\pi}{30} t) \\ p_2(t) &\sim 2sin(\frac{\pi}{30} t + \frac{\pi}{2}) \\ \end{aligned}$$
Plot the behavior of both populations on a graph, (...)
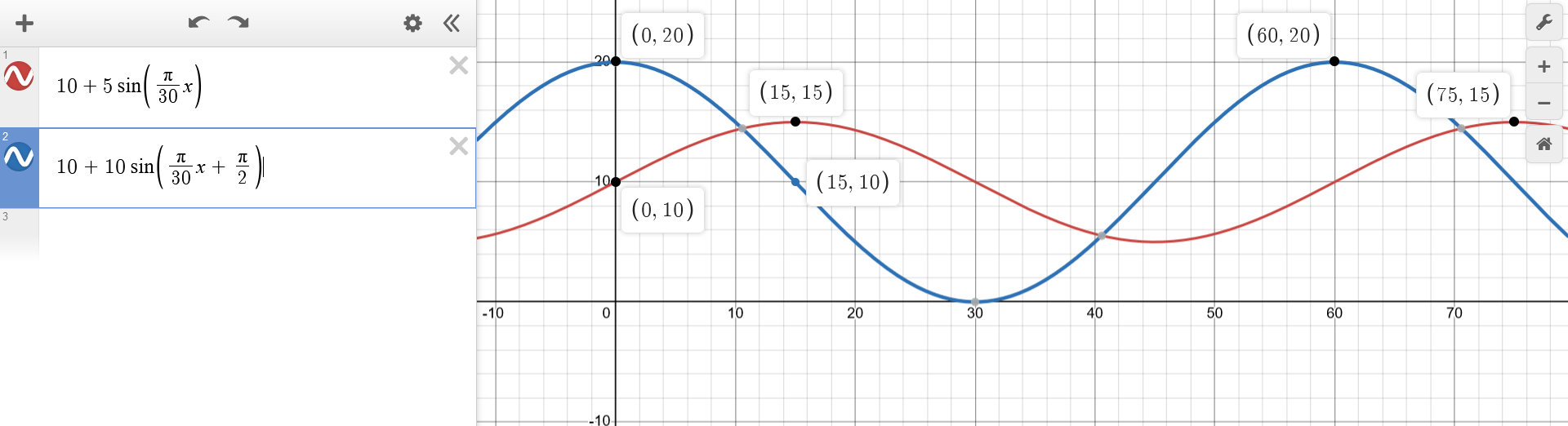
At what point in the cycle are the two populations equal?The plot shows that $p_1$ and $p_2$ intersect around $t \approx 10.5+30n$ for $n \in \mathbb{N}$. One can solve $p_1(t) = p_2(t)$ for a precise answer. $$\begin{aligned} p_1(t) &= p_2(t) \\ b + a * sin(\frac{\pi}{30} t) &= b + 2a * sin(\frac{\pi}{30} t + \frac{\pi}{2}) \\ sin(\frac{\pi}{30} t) &= 2sin(\frac{\pi}{30} t + \frac{\pi}{2}) \\ sin(\frac{\pi}{30} t) &= 2cos(\frac{\pi}{30} t) \\ tan(\frac{\pi}{30} t) &= 2 \\ \frac{\pi}{30} t &= arctan(2) + n\pi \\ t &= \frac{30}{\pi}(arctan(2) + n\pi) \\ t &= \frac{30}{\pi}arctan(2) + 30n \\ \end{aligned}$$ And indeed $t = \frac{30}{\pi}arctan(2) + 30n \approx 10.572 + 30n$.
When the prey population is at its maximum, what is the population of predators?The plot immediatly reveals that $p_2(t)$ has maxima at $t = 60n$ and that $p_1(60n) = b$. This can be confirmed by finding the $argmax$ of $p_2(t)$, given the known value of $argmax(sin) = 2\pi n + \frac{\pi}{2}$. Note that the $argmax$ is affected by function manipulations along the x-axis and unaffected by those along the y-axis. The former must be "undone" while the latter can be ignored. $$\begin{aligned} argmax(p_2(t)) &= argmax(b + 2a * sin(\frac{\pi}{30} t + \frac{\pi}{2})) && \\ &= argmax(sin(\frac{\pi}{30} t + \frac{\pi}{2})) && \text{ignore y-axis manipulations} \\ &= \frac{30}{\pi}(argmax(sin(t)) - \frac{\pi}{2}) && \text{"undo" x-axis manipulations} \\ &= \frac{30}{\pi}(2\pi n + \frac{\pi}{2} - \frac{\pi}{2}) && \text{replace known argmax(sin)} \\ &= \frac{30}{\pi}2\pi n && \\ &= 60n && \\ \\ p_1(60n) &= b + a * sin(\frac{\pi}{30} 60n) && \\ &= b + a * sin(2\pi n) && \\ &= b && \\ \end{aligned}$$
How does it change at that moment?The plot makes evident that $p_1(t)$ increases at $t = 60n$. The derivative $p_1'(t) = \frac{d}{dt} p_1(t)$ indicates how much. $$\begin{aligned} p_1'(t) &= \frac{d}{dt} [b + a * sin(\frac{\pi}{30} t)] \\ &= a*cos(\frac{\pi}{30} t) \frac{d}{dt} [\frac{\pi}{30} t] \\ &= \frac{a \pi}{30} cos(\frac{\pi}{30} t) \\ \\ p_1'(60n) &= \frac{a \pi}{30} cos(\frac{\pi}{30} 60n) \\ &= \frac{a \pi}{30} cos(2\pi n) \\ &= \frac{a \pi}{30} \\ \end{aligned}$$
And when the predator population is at its maximum, what is the population of prey?Again, the plot reveals that $p_1(t)$ has maxima at $t = 60n + 15$ and that $p_2(60n + 15) = b$ too. This can be confirmed by finding the $argmax$ of $p_1(t)$ in a similar manner as before. $$\begin{aligned} argmax(p_1(t)) &= argmax(b + a * sin(\frac{\pi}{30} t)) && \\ &= argmax(sin(\frac{\pi}{30} t)) && \text{ignore y-axis manipulations} \\ &= \frac{30}{\pi}(argmax(sin(t))) && \text{"undo" x-axis manipulations} \\ &= \frac{30}{\pi}(2\pi n + \frac{\pi}{2}) && \text{replace known argmax(sin)} \\ &= 60n + 15 && \\ \\ p_2(60n + 15) &= b + 2a * sin(\frac{\pi}{30} (60n + 15) + \frac{\pi}{2}) && \\ &= b + 2a * sin(2\pi n + \frac{\pi}{2} + \frac{\pi}{2}) && \\ &= b + 2a * sin(2\pi n + \pi) && \\ &= b && \\ \end{aligned}$$
How does it change at that moment?The plot makes evident that $p_2(t)$ decreases at $t = 60n + 15$. The derivative $p_2'(t) = \frac{d}{dt} p_2(t)$ indicates how much. $$\begin{aligned} p_2'(t) &= \frac{d}{dt} [b + 2a * sin(\frac{\pi}{30} t + \frac{\pi}{2})] \\ &= \frac{d}{dt} [b + 2a * cos(\frac{\pi}{30} t)] \\ &= -2a*sin(\frac{\pi}{30} t) \frac{d}{dt} [\frac{\pi}{30} t] \\ &= -\frac{a \pi}{15} sin(\frac{\pi}{30} t) \\ \\ p_2'(60n + 15) &= -\frac{a \pi}{15} sin(\frac{\pi}{30} (60n + 15)) \\ &= -\frac{a \pi}{15} sin(2\pi n + \frac{\pi}{2}) \\ &= -\frac{a \pi}{15} cos(2\pi n) \\ &= -\frac{a \pi}{15} \\ \end{aligned}$$